Blog | Reading Time 10 minutes
Fiber, an untapped energy source?
Turning fiber into milk: role of the rumen engine
Current context of high price of feed raw materials is driving us to reconsider the importance farm-grown fibrous material in the ration of ruminant. Indeed, fiber is an essential component of plants which is resistant to degradation by the digestive enzymes of most monogastric animals. Ruminants, on the other hand, have the unique ability to release the energy from part of the fiber structure thanks to the fermentative activity of the rumen. Ruminant digestive system has the ability to degrade and ferment fiber through microbial activity. The fiber components, and the subsequent production of volatile fatty acids (VFAs), provide the majority of the energy for the ruminant. Hence, to optimize ration to maximize fiber digestibility to extract more energy from the cell wall components of the plant. Better understanding of how fiber is degraded in the rumen helps identify the levers that can help reach this goal and make sure fiber is no longer an untapped energy source.
From fiber to energy to milk
Fiber is slowly degraded in the rumen due to its physical structure and the lignin cross-linking that holds the fiber intact. Fiber degradation in the rumen is influenced by:
- The anatomy of the fiber (pectin, hemicellulose and cellulose concentration), which is related to species
- Lignin content and structure (maturity, species, stressors, etc.)
- Particle size
- Passage rate (highly influenced by particle size, uNDF, DMI) and its impact on rumen dynamics
- Cud-chewing and ruminal contractions
- Rumen microbial population (microbiome) influenced by several factors (diet, environment)
The ruminant reduces the particle size of the forage through the eating process via initial mastication (chewing). These particles are swallowed and float to the top of the rumen mass. The particles are then regurgitated and rechewed (cud-chewing) to increase moisture content and surface area of the material as well as to abrade the fibrous portion.
After initial mechanical abrasion of fiber through mastication, the feed is then exposed to the diverse population of microorganisms in the rumen. The first step in fiber degradation is the colonization in the rumen of the particle of fiber by fungi and bacteria. Fibrolytic microbiota are very sensitive to oxygen. Strict anaerobic conditions (measured though redox potential) are necessary to ensure the maximum colonization. Many species of bacteria can produce these enzymes and are broadly categorized as fibrolytic bacteria. Furthermore, other microorganisms can similarly degrade fiber including protozoa and fungi.
The extent of ruminal fiber digestion is influenced by the digestibility of the fiber (NDFd – see boxed text), the rate of degradation of the fiber (Kd of NDFd) and the passage rate (Kp) of the fibrous material from the rumen.
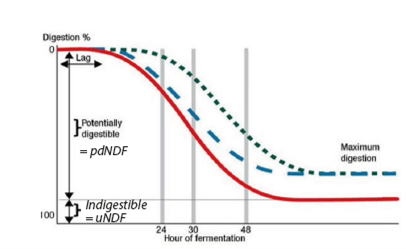
Figure 1: Fiber digestibility depending on rumen conditions and fiber type (MB Hall, 2014).
- The digestibility (NDFd) of the forage is influenced by morphological factors such as maturity (leaf:stem ratio) and lignification, plant anatomy and plant species.
- The rate of digestion (Kd of NDFd) is largely a function of species differences, lignin complexes and ratios, and maturity.
- The passage rate (Kp) can be influenced by theoretical length of cut, rumen dysfunction (acidosis and SARA), moisture content and dry matter intake of the animal.
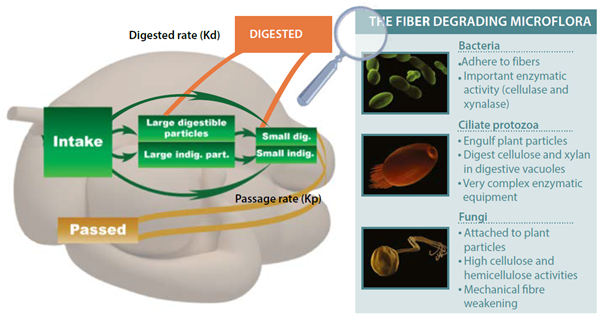
Figure 2: Fiber degradation in the rumen: kinetics and associated microbiota.
Fiber degradation in the rumen is leading to production of VFAs that represents up to 70% of the energy required for milk production. Thus, the efficacy of fiber degradation in the rumen is an important driver of milk production yield. Oba and Allen have determined that, for every 1% increase in NDF digestibility, milk production increases by 0.24 kg/d, and Fat Corrected Milk (FCM) increases by 0.25 kg/d (Oba and Allen, 1999).
Factors affecting fiber digestibility
There are several factors that can alter the population of the rumen microorganisms (rumen microbiome), including the influence of diet.
Ruminal pH and SARA
Since fibrolytic bacteria need a pH above 6.2, decreases in pH can inhibit their growth and performance. If rumen pH falls below 6.0, fiber digestion in the rumen begins to decline. When ruminal pH falls below 5.8 to 5.9, the rumen is mildly acidic, and fiber digestion in the rumen is disrupted.
Hence, any diet that can alter the pH can negatively impact fibrolytic species and result in reduced fiber degradation. This decreased pH can further lead to subacute rumen acidosis (SARA).
SARA can be influenced by a number of factors such as forage particle size, feeding behavior, starch content of the diet and grain level of the diet.
A meta-analysis by Ferraretto et al. (2012) examined factors that affected the digestibility of whole-plant corn silage. As starch levels increased in the diets, the digestibility of dietary NDF decreased by 0.61% units ruminally and experienced a 0.48% unit total-tract per %-unit increase in dietary starch content. The authors suggested this was due to a downward shift in ruminal pH.
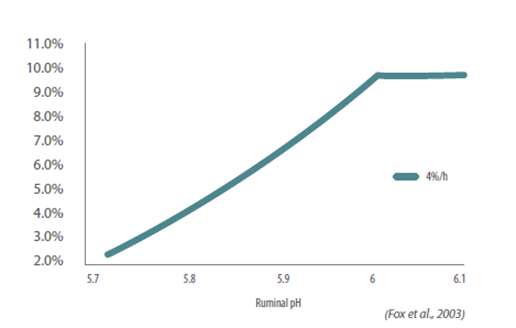
Figure 3: Impact of the rumen pH on the NDF digestion rate of a raw material (Fox et al., 2003).
Impact of particle size
The particle size of the ingested forage can play a role in fiber digestibility. Longer particles may increase a lag to digestion due to a lag in hydration and a reduction in bacterial attachment. Bacterial attachment refers to the ability of the bacteria to start the digestion process by attaching to the cut end of the fiber. The bacteria digest the plant material from the inside out and require either a cut point or opening created by fungal digestion or abrasion to start this digestion process.
Feeding behavior
Dairy cattle are foragers. Over the millennia, cattle have grazed on fibrous pastures. Ruminants have the capacity to consume human inedible feed, including the digestion of cellulose, with forages as the main source thanks to grazing and foraging.
The grazing/foraging behavior would be spread out over a long period of time with frequent small meals. Considering the modern dairy cow now has access to a total mixed ration (TMR), they typically consume their daily DMI in three to five hours per day, spread between 6 to 10 meals with the largest meal occurring after the delivery of fresh feed (DeVries et al., 2016; DeVries and von Keyserlingk, 2005).
Rumen pH generally declines after each meal as rumen available carbohydrates are fermented and volatile fatty acids (VFAs) are produced. When VFA production exceeds the ability of the rumen environment to neutralize or absorb the acids, SARA occurs. The size of the meal and the amount of rapidly fermentable carbohydrates can have an impact on the decline in rumen pH. Having smaller and more frequent, meals reduces variability in rumen fermentation patterns as well as the rate of pH decline and the amplitude of the rumen pH. In consideration of these factors, it is safe to conclude that changes in feeding behavior as a result from TMR feedings can increase the risk of SARA. Changing feeding behavior can reduce the risk of cows experiencing SARA.
Ruminants however have the biggest capacity to consume human inedible feed, including the digestion of cellulose, with forages as the main source but also feed materials such as sugar beet pulp, citrus pulp and brewers’ grains. Their unique ruminant digestive system allows them to convert “low value” fiber-rich feed components into “high value” animal proteins (milk and meat) not in competition with human foods, goes largely unnoticed.
Rumen modifiers
Rumen specific live yeast Saccharomyces cerevisiae CNCM I-1077 (LEVUCELL SC) used as a feed additive, has the ability to enhance fiber degradation through its rumen modifier effects, translated into higher feed efficiency in dairy cows.
The effects and modes of action of this live yeast on rumen microbiota have been extensively studied. The main effects attributable to this strain include:
- stabilization of ruminal pH
- increase in fiber degradation and the subsequent improvement in digestibility
In sacco, trials demonstrate the positive effect of S. cerevisiae CNCM I-1077 on NDFd of more than 200 different forage samples, including: corn silage, straw, rye grass hay, alfalfa hay, annual rye grass (pasture), meadow hay, grass silage, etc. Saccharomyces cerevisiae CNCM I-1077 is found to increase NDF digestibility by 4 to 8 units, depending on the type of forage and on its own degradability (Guedes et al., 2008; Guedes et al., 2015; Chaucheyras-Durand et al, 2010, Ding et al, 2014).
Research studies have evaluated the possibility to model the effect of S. cerevisiae CNCM I-1077 on dynamic model. It clearly demonstrates that the effect of a rumen modifier based on its mode of action while well documented, can be integrated into a nutritional software and accurately predict the effect on animal performance.
Overall, combined data trials indicate a 3% improvement of feed efficiency with the live yeast supplementation, with higher effect under rumen challenged conditions (up to 7% increased feed efficiency, corresponding to a 6:1 up to 9:1 return on investment for the producer under these challenging situations (figure 4).
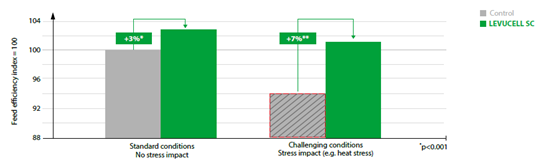
Figure 4: Levucell SC effects of dairy cow feed efficiency (average of several trials).
Fiber represents the majority of milk energy for dairy cows. Fiber degradability is linked:
- to the quality of the plant material, which depends on both the crop inherent characteristics and its growing environment, and
- to the fiber digestion process once it is fed to the animal (mainly located in the rumen).
So, the degradation of fiber is influenced by farming practices and rumen dynamics. Indeed, the rumen can be pictured as an engine that, thanks to its unique microbiome, turns fibers into milk energy. Management and feeding practices that ensure optimal rumen environment (pH, microbial balance…) and function can help that improve the efficiency of the ‘rumen engine’ for optimal farm revenue.
Learn more about Levucell SC!
References:
-
Chaucheyras-Durand F., A. Ameilbonne, N. D. Walker, P. Mosoni and E. Forano. 2010. Effect of a live yeast, Saccharomyces cerevisiae I-1077 on in situ ruminal degradation of alfalfa hay and fiber-associated microbes. J. Anim. Sci. 88 (S2)/J. Dairy Sci.93 (S1): 145
-
DeVries T. J. 2016. Something to Ruminate on…Why We Should Watch Cows Chew. WCDS Advances in Dairy Technology. 28: 99-108.
-
DeVries, T. J., M. A. G. von Keyserlingk, and K. A. Beauchemin. Frequency of Feed. Delivery Affects the Behavior of Lactating Dairy Cows, J. Dairy Sci. 88:3553–3562.
-
Dhiman, T.R., M.S. Zaman, I.S. MacQueen, and R.L. Boman. 2002. Influence of Corn. Processing and Frequency of Feeding on Cow Performance. J. Dairy Sci. 85:217–
-
Ding G., Y. Chang, L. Zhao, Z. Zhou, L. Ren and Q. Meng. 2014. Effect of Saccharomyces cerevisiae on alfalfa nutrient degradation characteristics and rumen microbialpopulations of steers fed diets with different concentrate-to-forage ratios. J. Anim.Sci. and Biotech. 5: 24
-
Ferraretto, L.F. and R. D. Shaver. 2012. Meta-analysis: Effect of corn silage harvest practices on intake, digestion, and milk production by dairy cows. The Professional Animal Scientist. 28:141-149.
-
Fox D. G., T. P. Tylutki, L. O. Tedeschi, M. E. Van Amburgh, L. E. Chase, A. N. Pell, T. R. Overton, and J. B. Russell. 2003. The Net Carbohydrate and Protein System for evaluating herd nutrition and nutrient excretion. CNCPS version 5.0. July 29, 2003.
-
Guedes C. M., Gonçalves D., M. A. M. Rodrigues M. A. M. and A. Dias-da-Silva. Effect of a Saccharomyces cerevisiae yeast on ruminal fermentation and fibre degradation of maize silages in cows. Anim. Feed Sci. Technol. 145: 27-40
-
Oba M. and M. S. Allen. 1999. Evaluation of the importance of the digestibility of neutral detergent fiber from forage: effects on dry matter intake and milk yield of dairy cows. J Dairy Sci. 82(3):589-96.
Published Jan 17, 2023 | Updated May 29, 2023
Related articles
Need specific information?
Talk to an expert


